始めに / 品種に就いて / 遺伝概論 / 品種一覧 / 付箋集 / 参考文献 / 飼育に就いて / 諸問題と解決方法 / 繁殖に就いて / 終わりに
遺伝概論
◇単語/words
◇遺伝子/gene
◇染色体/chromosome
◇遺伝子座/locus
◇表現型/phenotype
◇対立遺伝子/allele
◇遺伝子型/genotype
◇接合/junction
◇突然変異/mutation
◇野生型/wild type
◇優性と劣性/dominant and recessive
◇メンデルの遺伝法則/Mendel's law of heredity
◇パンネットのスクエア
◇複対立遺伝/multiple allelomorphs
◇共優勢遺伝/co-dominant inheritance
◇優性遺伝/dominant inheritance
◇その他の遺伝
◇細胞質遺伝/cytoplasmic inheritance
◇伴性遺伝/sex-linked inheritance
◇致死遺伝子/lethal gene
◇同義遺伝子/multiple gene
◇抑制遺伝子/inhibitor gene
◇相加遺伝子/additive gene
◇多因子遺伝/multifactorial genetic
 遺伝概論
遺伝概論
遺伝に就いて。
此処から下には管理人の文章構成能力の無さ故に難しげになってしまっているけれども、書いてある内容は至って易しい文章が書き連ねてあるわけですが、斯うした遺伝子の種類や遺伝の法則を理解するのは、何のためにあるのか、ということを最初に。
遺伝を学ぶ事による有用性一つ一つを理由として挙げるのは容易いのですが、それはやめておきましょう。それらに於ける価値は、十人十色でありましょうから。………ブリーダーであるなら、多重劣性遺伝表現型などの作出を効率的に行うにあたって有用であるから、という言い方も出来るでしょうし、ただ単純に、今いる個体と同じような品種を作りたいのだけれど、それにはどうすれば?という疑問を理由とするのでもよいでしょう。なんであってもよいと思います。
故に、強いて此処で一つの見解を示すとすれば、それは管理人個人の見解となるでしょう。その見解にしても、そんな大した事でもないのですが。
管理人自身は、こう思っています―――現在、様々な遺伝性の変異の組み合わせにより沢山の品種が確立している類のヘビにあっては、遺伝の面白さを理解すると、楽しみ方の幅が一気に広がるだろう、と。
繁殖をしないで、ただ飼育するだけが趣味の方もいるかと思います。そういう場合には、遺伝の知識は必要ないと言えばないでしょう。然し、現在飼育している個体の起源を辿り、それが作出されるまでの歴史を振り返り、作出方法を理解していくことで、より思い入れが深くなるかもしれない。……まぁ、ならないかもしれませんが(苦笑)
とまぁ、管理人がぐだぐだ言わずとも、皆様は今、この文章を読んでいるのですから、遺伝にちょっと興味が湧いているという事なんだと思います。
此から続く文章は、慥かに最初は取っ付き辛いかと思いますが、言うなれば”役”のあるゲームのようなもので、役の材料となる札(=遺伝子)がどんなものかを覚えて、それを組み合わせる事の出来るルール(法則)を把握し、不確定にして不条理な賽子(さいころ)の目に達観しましょう、ってことが書いてあるにすぎません。管理人が思うに、麻雀やトランプのナポレオンよりはずっと簡単に覚えられると思いますよ。
え? 麻雀もナポレオンも、どっちもできない? そうですか、それは困りましたね………………う~ん、でも、管理人もどっちも出来ませんから、たぶん問題ないんじゃないでしょうか(笑)
最後に付け加えておくと、このページの遺伝は、管理人が色々集めて読んだ記憶や知識を頼りに、蛇の突然変異による品種の説明をより分かり易くするために構築した文章であって、遺伝学の文章ではありません。遺伝に就いて、ちょっと古いことが書いてあるかもしれません。そのほうが分かり易いと思えば、古い書き方を採用したからです。また、説明しなくてもどうにかなってしまうことは省いてあったりします。
とはいえ、間違いは原則的に無いとは思うのですが………もし、何処か妙なところがあったら、メール等で指摘していただけると有り難いです。
それでは、比喩ではなく正しく”目眩く”遺伝の世界へ、漕ぎ出すと致しましょう~
単語/words
 遺伝子/gene
遺伝子/gene
遺伝子とは、DNA(=デオキシリボ核酸)の中にある、情報伝達の”単位”の事。
DNAには、その生物の体の構築、生命活動に必要な様々な蛋白質の設計図など、生命の発生から成長、生命活動に必要となるあらゆることが書かれていると言われる。遺伝子というのは、DNAの上に塩基配列で書かれた数々の設計図、その部分部分の名称と考えればよい。塩基一つ一つが遺伝子なのではなく、DNAの此処から此処までの部分が、Aに関わる設計図になっているとき、その部分が、遺伝子Aと呼ばれる事になる。厳密な意味で遺伝子地図が解析されている例は少ない。
◇染色体/chromosome
DNAは、細胞の細胞核に存在するが、細長い構造をしているため、内部に収納するにあたって、折りたたむ必要がある。DNAにある種の蛋白質が巻き付き、小さな構造を構築し、さらにそれらが巻き合って大きな構造――染色体を形成するのだという。染色体の形としてもっとも一般的に想起される形態は、細胞分裂時に出現するX型のものと思われるが、此は細胞の中に二対一組の遺伝子を持つ二倍体の生物であるからこのような形になるのであり、すべての生物の細胞が、細胞分裂時にああした形態をとる訳ではない。
端的に言えば、単為生殖する種の細胞には一セット(x本)の染色体しかないのに対し、有性生殖する種の細胞には、父親と母親から受け継がれた染色体がそれぞれ一セットずつ、合計二セット(2x)の染色体が存在する。二セットの染色体が存在する細胞を二倍体と言い、三セット以上もつ細胞を多倍体(nx)と言う。
有性生殖を行う種では、体細胞は二倍体(2x) かまたは多倍体(nx)、配偶子は半数体(x)になる。(ex.人間の肝細胞は一つの細胞の中に核が二つあることが知られる。よって、人間の場合でも、総ての体細胞が二倍体だという訳ではない)。
人間の染色体は一セット(x=)23本あり、細胞には両親からそれぞれ受け継がれた染色体が対になって合計46本存在する。対になっているというのは、対になった染色体同士は、内容は異なるが、同じ内容を司る遺伝の配列になっているという事。これを相同性と云う。例外は、人間の場合、23番目のナンバーが振られた染色体で、此は性染色体と呼ばれ、X染色体とY染色体という構造が異なるもので、対になってはいるが、二つの染色体に相同性はない為、例外視されている。
此の性染色体を除く染色体の対は、全長に渡り配列がこの相同性を持っていることから、相同染色体と呼ばれる。性染色体に対し此らは常染色体と呼ばれているが、単為生殖する生物などではあまり意味がない概念でもあるらしい。
性染色体はヒトの場合X,Yの種類で呼ばれるが、鳥類や爬虫類などではZ,Wの組み合わせが使われている。ヘビの染色体数は種によって異なるが、日本のシマヘビでは38本。ヒトがXYが雄、XXが雌になるのに対し、ヘビなどの爬虫類は、Zを2本持つ方が雄になり、W染色体を持つ方が(つまりWWとWZが)雌になるとされる。
常染色体――対となる相同染色体が持つのは相同性であり、同一性ではない。配列は類似性を持つが、それは、同じ種類の遺伝子を持っているという事で、同一の遺伝情報を持っている訳ではない。
常染色体の対となる染色体の一本一本(染色分体)は、それぞれ、その母親と父親から一本ずつ受け継がれるものであることが知られている。
何故、一本ずつ受け継がれるのかと言えば、生殖細胞である配偶子(精子と卵子)が持つ染色体が、一本だけだからに他ならない。
通常、細胞(真核細胞)が分裂するとき、染色体の数が減らないよう、染色体の複製が行われ(核分裂と言う)、染色体を二倍持つ四倍体と呼ばれる細胞になる。
そしてそこから細胞分裂が起こり、染色体は半分ずつそれぞれの細胞に受け継がれ、二つに分かれた細胞はそれぞれ、もとの細胞と同じ量、そして複製なので同じ内容の染色体を持つことになる(厳密にはテロメアが異なるが、此処では省略する)。
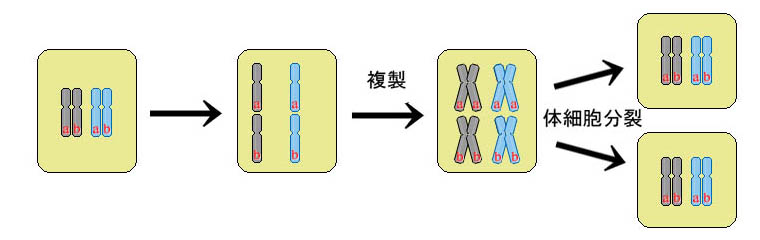
もしも、配偶子を作る時にも、同じ様に分裂した場合、配偶子が持つ染色体の数は、ヒトの場合で体細胞と同じ本数であるから、46本になり、その配偶子同士が受精した場合、染色体の数が92本になってしまう。
現実にはそんなことは起こりえないのだが、それは、配偶子を作る際に行われている分裂が、体細胞分裂とは異なる減数分裂と呼ばれる分裂だからである。
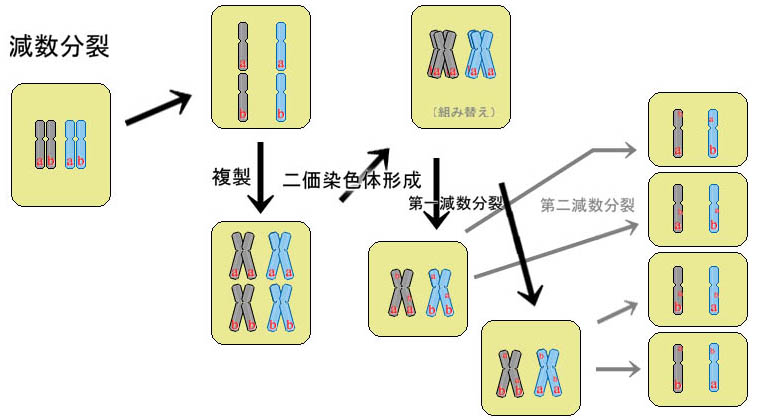
減数分裂とは、配偶子(精子や卵子)が作られる時のみに行われる分裂で、一つの細胞が最終的に四つに分裂するが、染色体の複製が一度しか起こらない。結果、細胞の中の染色体の数が最終的に半分になることから、減数分裂、と呼ばれる。
過程としては、最初は体細胞分裂とほぼ同じで、細胞は染色体を内部で複製する。此により染色体の数(DNAの量)が二倍になる。
此の状態で、分裂が起こらず、相同染色体同士が近接し、二価染色体と呼ばれるものを形成する。此の二価染色体では組み替え(乗り換え)と呼ばれる現象が起こる。
組み替えとは、其の個体が両親から受け継いだ染色分体がお互いに抱き合い、部分部分で情報の乗り換えが行われることで、分かれた時に、元から存在していた染色分体と全く異なった染色分体が形成される現象。此により、単純に両親から受け継いだ染色分体が、そのまま配偶子に組み込まれるという事は起こりえなくなっており、遺伝子の多様性を生み出す原理の一つと言われている。
その後、相同染色体(組み替え後)は離れ、分裂することで、第一分裂が終了する。この時、異なる相同染色体同士は別々に独立してランダムで分裂する細胞のどちらかに入る。
此処までで、通常と同じ量の染色体を持つ二つの細胞が出来るのだが(但し、染色体の情報は組み替えにより異なったものである)、此処で更に第二分裂が起こり、相同染色体は二本の染色分体となり、二つの細胞のそれぞれに分かれる。結果、最終的に出来た四つの細胞には、元の細胞の半分の量の、但し異なった遺伝情報が含まれる事になる。
此により、母親から一本、父親から一本の染色分体が継承され、様々な一つの染色体を持つ個体が生まれる様になっている。
遺伝子座/locus
遺伝子座とは、染色体や、DNAなどの上の、遺伝子が存在する位置の事。遺伝子地図や、此らの遺伝子座の距離により連鎖が起こるので、場合によっては重要になることもあるが、今回のヘビの遺伝の話に於いてはあまり必要がないので、その辺の話は省略。
かみ砕いて言えば、DNAの上の、特定の遺伝子が存在する場所。遺伝子が存在する場所があり、その場所に存在出来る遺伝子が複数である場合、その遺伝子の座の奪い合いになる訳だが、遺伝子座は、染色分体につき一つ存在するので、通常、席は二つあることになる。
若しも、一つの遺伝子座に於いて、三つや四つ以上の遺伝子が存在する場合でも、一つの個体は二つしか遺伝子を持ち得ないので、一つの座につき、最大で二つ以上の遺伝子の保有者にはならない。
表現型/phenotype
生物が持つ、生理的な形質や性質、形態や機能の中でも、遺伝的に継承されるものを言う。個体が生存していく過程で獲得したような形質は含まれない。例えば、皮膚が硬質化して形成されたと言われる人間の爪が存在することは遺伝的なものであり、表現型と言えるが、同じ皮膚の硬質化でもペンだこのようなものは後天的な獲得形質であり、故に遺伝性がないため、表現型には含まれない。
表現型には、幾つかの要素が重なり合ったものや、一つの遺伝子の変異で簡単に支配されてしまうものなど色々ある。
単純に支配される要因で左右される表現型もあれば、複数の同様の機能を持つ遺伝子が存在することで(同義遺伝子)、一つぐらいの変異では表現型にならないものや、性染色体のどちらかにしか継承されないで、その上変異を継承している遺伝子が不活性化している場合などは表現型にならない(後述アリ)
ハープタイルホビーの分野では、或る何かしらの遺伝性突然変異形質が重要視されるが、個体群もある種の表現型である。
但し、個体群という一つの表現型は、多因子によるものであり、単純に遺伝するものではない。その上、通常、形質にばらつきがある。このばらつきを浸透度と言うが、浸透度は、方向性を揃える事で度数を変化させることが出来る。此の発想が選抜交配であり、選別と選抜を重ねに重ねると浸透度が深くなり、或る一定レベルを超えると、ばらつきが極めて少なくなる。ばらつきが少なくなったならば、それは品種として完成したと言えるのだろう。
此の最たる例がブラッドレッドで、ブラッドレッドの腹部の白(プレーンベリー)は単純に劣性遺伝するという(優性遺伝の可能性もアリ)。しかし、他の細胞分布や赤色色素増幅などは単純には遺伝しない。ブラッドレッドは、ある表現型を持つ個体群の浸透度を、人為的に高めた結果生まれた品種であると言える。
対立遺伝子/allele
相同な遺伝子座を占める遺伝子の種類が複数ある場合、これらを互いに対立することから、対立遺伝子と呼ぶ。
対立する遺伝子は、通常、突然変異によって生じると言われている。つまり、突然変異の遺伝子は、総て、野生型/wild typeに対して対立である。但し、どの遺伝子座でも野生型 /wild typeという言葉が使われるが、当然、赤色色素の生成に関わる遺伝子座と、模様に関わる遺伝子座に占める野生型
/wild typeの遺伝子は異なる遺伝子である。
遺伝子座に占める遺伝子が、野生型 /wild typeを含めて二つではなく、三つ以上存在する場合、複対立遺伝が起こる。詳しくは、後述「複対立遺伝」の項目を参照のこと。
遺伝子型/genotype
広義の遺伝子型とは、その個体が持っている総ての遺伝子の構造を云う。だが、此は広義すぎる言葉で、一般的には、対立遺伝子が存在する遺伝子座が、それぞれどのような遺伝子(遺伝子の変異)の組み合わせになって存在しているかを意味し、通常は特に記すべき遺伝子(その学問や研究に関連し重要な意味を持つ遺伝子)を表記する。突然変異に限らず、地域変異による遺伝子の違いなども、遺伝子の場所がどのような型になっているかで判別される(ウイルスの型など)。
表記方式はあるのだけれども、ハープタイルホビーの分野では、特に決まった書き方はないようで、人によって書き方が違う。とはいえ、結構分かるようになっているので問題はない。色彩変異、模様変異の遺伝子が重要なので、書かれるのは各色素変異及び模様変異になる。
接合/junction
ある遺伝形質を決定する遺伝子座があり、其処に収まる遺伝子には、Aとaという対立する遺伝子が存在する場合。
細胞が二倍体のとき、相同染色体の遺伝子座は、AA、Aa、aaという三つの形をとることになる。この時、AA、又は、aaという同一の遺伝子が遺伝子座を共に占めている状態を持つものをホモ接合体(homo zygote)と呼び、Aaという異なる対立遺伝子が座を占めている状態をもつものを、ヘテロ接合体(hetero zygote)と云う。
homoとは同種という意味であり、heteroとは異種という意味である。
突然変異/mutation
突然変異とは、遺伝情報に永久的な変化が起こることを言う。どの細胞でも起こりうる現象であるが、遺伝するには生殖細胞に変異が起こらないと意味がない。突然変異が起こるには様々な要因があり、紫外線や放射線は直接的な原因に成りうるし、化学的要因もあり得る。
ハープタイル・ホビーの分野では、目に見える色素、模様などの変異を特にさしてミュータントと言う。黒色色素、赤色色素、黄色色素などの種々の色素の生成に関わる遺伝子が変異を起こしたものや、体表面の模様の形成に関わる遺伝子が変異したものなどが、繁殖され維持されている。
此らの突然変異遺伝子は少なくとも野生型/wild type(正常な機能を持つもの)に対して対立である。
突然変異には、大きく分けて機能欠失型、機能獲得型がある。
前者の場合、機能を完全に失うとamorphと呼ばれ、部分的に失うとhypomorphになる。例えば、melanineに関わる遺伝子が、メラニン生成能力を失った場合、その遺伝子はAmelanistic遺伝子になり、部分的に失った場合はHypomelanistic遺伝子になる。
後者の獲得型は、野生型/wild typeの遺伝子の効果を”妨げる働きを持つ”antimorphと、全く異なる新しい機能を獲得するneomorphがある。通常、antimorphは、野生型に対して優性であり、amorphやhypomorphは劣性である事が多いようだ。
野生型/wild type
様々な突然変異(ミュータント)に対し、もともとの遺伝子の状態を野生型/wild typeと言う。長い間に培われたもので、通常様々な突然変異に対して優性に遺伝する。数千、数万年という長い目で見た場合、突然変異の変化を取り入れ、野生型も常に変化していくのであり、個体群によっては何が野生型か、というのが当然変わってくる。
優性と劣性/dominant and recessive
一つの遺伝子座に於いて、対立遺伝子がある場合、その二つの遺伝子の間に、表現型としての現れ方に優劣がある場合、一方が一方にに対し、優性である、或いは劣性である、という表現が為される。
通常、遺伝子は二つで一組となっているが、座に一つでも存在すれば表現型になる遺伝子は、その他の対立遺伝子に対して優性である、ということになる。此の場合、優性の遺伝子は座に一つ(ヘテロ接合体)のときも、二つの(ホモ接合体)のときも同じ表現型になる。若しもヘテロ接合体とホモ接合体とで表現型が異なる場合、その遺伝子は、ヘテロ接合体となっている相手の遺伝子に対して優性であると言えないので、優性遺伝子とは言えなくなる。
例えば、対立する遺伝子Aとaに於いて、Aが優性であり、aが劣性である場合、接合がAA、Aaの場合に表現型はAになり、接合がaaの場合にのみ、表現型はaになる。
優性、劣性は遺伝的性質のみに於いてのことで、遺伝子の機能には関係はない。日本語では、優性と劣性という呼び方になっているが、dominanceはもともと、支配するという意味の単語。
劣性遺伝子というのは、其れに対立する遺伝子があり、かつその遺伝子よりも優性である遺伝子がそこに存在するときに、相対的に規定されるものになる。よって、一つしか存在しない遺伝子座にあっては、優性遺伝子も劣性遺伝子も存在しない。
そうした遺伝子が存在するのかどうかは分からないが、例えば、一つの座に、遺伝子A、遺伝子B、遺伝子Cという三つの遺伝子が存在しており、遺伝子Aは遺伝子Bと遺伝子Cに対して優性であり、かつ、遺伝子Bは遺伝子Cに優性であるという場合も有り得るだろう。
そのとき、遺伝子Bは、「遺伝子Aに対して劣性であり、遺伝子Cに対しては優性」ということになる。よって、遺伝子Bをして優性遺伝子、であるとか、劣性遺伝子である、というような表現はありえなくなる。
○○が優性である、或いは、劣性である、という表現は、常に対立する遺伝子との相対的な関係を意味している。一般的にそうした部分を考えなくても意思疎通に障碍がないのは、特にハープタイルホビーの分野では、「野生型に対して、○○である」という意味合いで使うという前提になっていることと、上に挙げたような、三段階で優性劣性になっている遺伝子が存在しないからだろう。
 メンデルの遺伝法則/Mendel's law of heredity
メンデルの遺伝法則/Mendel's law of heredity
メンデルの遺伝の法則とは、Gregor Johann Mendel/グレゴール・ヨハン・メンデル(1822-1884年。グレゴールは洗礼名)によって1865年に出された「植物雑種に関する研究」に述べられていたもので、支配の法則(law of dominance)、独立の法則(law of independence)、分離の法則 (law of segregation) の三つからなる。
但し、メンデル自体はその論文の中で、斯うした名称を付けてはおらず、法則という名称も使わなかったようだが、法則に従っているとは思っていたようだ。故に、法則と呼ぶべきか呼ぶべきではないかは難しいところであると言える。
今日のメンデルの法則という呼称は、コレンス/C. Correns、チェルマク/E. Tshermak、ド=フリース/H. de Vriesという三人の学者が1900年に、この遺伝法則を再発見し、コレンスがメンデルの論文の根本原理をメンデルの法則と呼んだ事に起因するらしい。ただ、コレンスらはどうやら、上記三つの法則をしてメンデルの法則だと呼んだ訳ではないようであるが(此について述べた文章を読んでそのように判断したが、論文そのものを見ていないので断言しかねる)。それが何故に現在では、特に上記の三つをしてメンデルの法則と呼ぶ向きが定着しているのかまでは知らないが、おそらく、説明し易く、分かり易いからだろう。
但し、支配の法則(優性の法則、或いは優劣の法則)自体はメンデルが見出したものではないし、独立の法則自体も彼が見出したとはやや言い難い。彼最大の発見は、やはり分離の法則に尽きるとする意見もある。
何故かと言えば、彼が研究を書き記した当時は、染色体の存在も、減数分裂という現象も知られておらず、子が親に似るという、遺伝の現象を説明する理論として、経験的な観測から、親の精子と卵子に存在する液状の何かが存在し、それらが混ざり合うことで、両親の性質が子供に引き継がれると考えられていた。
メンデルは、其れに対し、遺伝は何かしら液体的なものではなく、単位化された粒子的な物質であることを予言したのであり、その論拠となるのが分離の法則だった訳である(此を粒子説と呼び、対して前述したような液状の何かが混ざることで遺伝が行われるという考えを、融合説、或いは混合説と現在では呼んでいる)。
では、どのような実験と思考の過程を経て、彼はそうした結論に達したのか。
メンデルの研究は非常に根気のいる作業から始まったようである。まず、エンドウ豆に、背の高いものと低いものがあることに彼は着目し、純系の選抜を行った。すなわち、背の高いものの種子だけを集め、離れた別の場所で育て、育ったものの中から、高くなったもののみの種子をまた集める。此を、必ず背が高くなるエンドウ豆の種子を収穫出来るようになるまで、数年繰り返した。
低いエンドウ豆も同様に別の場所で選抜を行った結果、必ず低くなる種子を収穫することが出来るようになったという。
メンデルは、必ず背の高くなるエンドウ豆の種子を育て、咲いた花のめしべ(雌蕊)に、必ず背の低くなるエンドウ豆の種子を育てて咲いた花の花粉を受粉させた。
逆に、背の低くなるエンドウ豆の方に咲いた華にも、背の高くなるエンドウ豆の花粉を受粉させた。
此ら二つケースで収穫されたエンドウ豆を、翌年播いて育てたところ、すべてのエンドウ豆の背が高くなった。
この総て背が高くなったエンドウ豆から収穫された種子を翌年撒いた。すると、背の高いものが全体の3/4であり、低いものが1/4になったという。
メンデルは、高さと低さといった性質だけではなく、エンドウ豆の種子に皺のあるものとないもの、種子の色が黄色と緑色のものなど、幾つかの形質に着目し、同じ様な実験を行った結果、しわのあるものと、ないものを交配すると翌年はしわのないもののみが収穫され、この種子を翌年育てると、しわのないものが3/4、しわのあるものが1/4収穫されるという結果を得た。
此処までは既存の混合説で説明が着くのだが、雑種二代目では、雑種一代目では隠れていた劣性の性質が表面に現れるとういう現象は、両親は共に、子に現れる性質を持っていないのに、子には引き継がれているのだから、説明出来ない。
メンデルは此らの実験結果を、まず、両親から子へは単位化された粒子(要素と呼んだ)による何かで遺伝の性質は引き継がれ、それらの性質を持つ粒子には、優先されるものと、優先される見かけ上は隠れるものがあるとした。(優性の法則。もともとは、その性質を支配する、という意味合いから、支配/dominanceと呼んだ。これが日本で何故優性という言葉になったのかは分からない)
そして、雑種二代目にしてその性質が分離して現れる事、その出現する確率現象から、彼は個体は、雌性の配偶子と雄性の配偶子(この場合は花粉)に由来する一対の”要素”をもち、かつ、個体が配偶子をつくるときに、この対になった要素一つずつに分かれて別々の配偶子に入ると結論付けた。
すなわち、エンドウ豆が背の高いものと低いものの雑種一代目がすべて背が高くなるのは、背が高い方が、低い方に対して支配的であるからである。そして、両親から半分ずつに受け継がれた要素は、次の代に引き継がれる時に、半分ずつに分かれ、半分の確率で遺伝する。
高さの要素Hは、高い(H)と低い(L)があるとき、
H(H.H)×H(L.L)=H(H.L)
となり、Hの方がLよりも支配的であるので、すべてのエンドウ豆株は高くなる。雑種二代目は、
H(H.L)×H(H.L)=H(H.H)1/4,H(H.L)1/4,H(L.H)1/4,H(L.L)1/4
H(H.L)はすべての株が高くなるので、高くなる株と低くなる株H(L.L)の比率が、3:1になるのだ、という訳である。仕組みを知る我々には納得であるが、現象から此らの事を洞察出来たのは、彼の洞察力が優れていたこともあろうが、なによりも、非常に分かり易い形質に絞ったこと、最初の数年を純系の系統の確立から始めたこと、詳細なデータを蓄積し、統計的に解析したことなど、総合的で論理的な考察に基づいているからだと云われている(ただ、理論に沿うように恣意的に観察してしまったという意見もあり、実際、その為に起こったと思われる誤謬も指摘されている。それらの事はこの頁で述べる必要がないので省略する)。
メンデルは複数の異なった形質に於いて遺伝の仕方を調べた事は前述したが、彼はそれらの形質が、それぞれ干渉しないことを述べている。つまり、エンドウ豆の株の高さの要素と、種子の皺の要素は別々に遺伝しており、それぞれがどういう表現型であるかによって、もう一方の表現型が変化するようなことはないとした。此が、独立の法則と呼ばれるものになる。
教育論などに目を向けると、メンデルの法則は正しいだとか正しくないだとか云う言説が少なからずあり、此を書くにあたって少々困惑したが、なんとなくざっとそれらの主張を読んでみて、気にしないで良さそうだと思ったので、上のような感じになっている。まぁ、皆様も別に気にする事はないかと。
パンネットのスクエア/Punnett square
格子分析とも云われる此の手法は、欄外の縦と横に配偶子に成りうる要素に、その出現確率を書き添えたものを並べ、格子(菱形、或い四角)の中に、接合子と、その出現確率(此は、組み合わさる配偶子の出現確率の乗になる)を書き添える。
スクエアの中の合計は百分率で100になるか、或いは分数表記の場合は1になる。各格子に書き込まれる確率は、総てが分数だと1/配偶子の種類の二乗、になる為、省略されることもあるが、直視的に理解しやすい事もあり、省略しないほうが望ましいとされているという。当サイトでは省略しないことにする。
余談だが、Punnettの発音は表記するとパネットに限りなく近いようで、そう書かれる事もあるが、此処ではパンネットのスクエアとした。スクウェアかスクエアかというのも、同じように難しいところなのだが、此処では、なんとなく管理人の趣味でパンネット・スクエアに決めることにした。呼びたい人は、パネットのスクエアでも、パンネットのスクウェアでもお気に召すまま気の向く儘に。
Melanine (n=normal, a=amelanistic) |
M(n) 50% | M(a) 50% |
| M(n) 50% | M(nn) 25% | M(na) 25% |
| M(a) 50% | M(na) 25% | M(aa) 25% |
上記は雄、雌共に、M(n,a,)という接合の個体であるケース。配偶子(精子と卵子)は、50%の確率でnとaの因子のものになるので、その組み合わせは、それぞれの確率を掛けた数字となる。
Melanine (n=normal, a=amelanistic) Hypo (n=normal, h=hypomelanistic) |
M(n)H(n) 1/4 | M(n)H(h) 1/4 | M(a)H(n) 1/4 | M(a)H(h) 1/4 |
| M(n)H(n) 1/4 | M(nn)H(nn) 1/16 | M(nn)H(nh) 1/16 | M(na)H(nn) 1/16 | M(na)H(nh) 1/16 |
| M(n)H(h) 1/4 | M(nn)H(nh) 1/16 | M(nn)H(hh) 1/16 | M(na)H(nh) 1/16 | M(na)H(hh) 1/16 |
| M(a)H(n) 1/4 | M(na)H(nn) 1/16 | M(na)H(nh) 1/16 | M(aa)H(nn) 1/16 | M(aa)H(nh) 1/16 |
| M(a)H(h) 1/4 | M(na)H(nh) 1/16 | M(na)H(hh) 1/16 | M(aa)H(nh) 1/16 | M(aa)H(hh) 1/16 |
上記は、共にアメラニスティックとハイポメラニスティックの因子を、M(na)H(nh)という形で持つ個体同士を掛け合わせた場合の格子になる。確率は25%でもよいが、小数点以下が続くと煩雑になるので、分数の方が分かり易いかもしれない。
このパンネットのスクエアを使うと、視覚的にメンデルの法則の骨子を理解しやすく、義務教育課程の何処かで、誰でも一度は見たことがあると思われる。メンデル自身は此の様な書き方はしなかったようで、数式のように並べていたらしい。だからこそ、此はパンネットのスクエアであって、メンデルのスクエアではない。
複対立遺伝/multiple allelomorphs
遺伝学の初期は、個々の座に於いて確認されていた対立遺伝子の数が少なかった事から、一つの座に於いて複数の対立遺伝子が存在する遺伝方式を複対立遺伝と呼んだが、遺伝解析が進んだ後、理論上、遺伝子の最小単位は微小であるが故に対立遺伝子は非常に数多く存在出来ることが分かり、実際一つの遺伝子座に対し多くの遺伝子が見つかったことから、敢えて名称を付ける必要性がなくなり、複対立という呼称は形骸化した。例えば、一つの座に対して1000以上の対立遺伝子が確認されている例もある。
但し、ハープタイルホビーの分野では”目に見えて意味のある”突然変異にのみ限った話なので、そうなると種類は数えられる程であり、この概念も有用であると云え、また理解のし易さからも使用されている。
敢えて複対立遺伝という言葉を使う理由は上に述べたが、では複対立遺伝とは如何なるものかを再確認しておくと、ある座に於いて、三つ以上の対立遺伝子が、その種若しくは個体群内に存在する場合に、その遺伝子座は複対立遺伝子を持つ、と表現される。種、若しくは個体群内に、という条件を付けたのは、爬虫類や哺乳類など、二倍体の生物の体細胞は、ある特定の座に於いて、正常な範囲では二つのみ遺伝子を持つものであり、三つ以上の遺伝子を内包することは有り得ないからである。
また、この際、複数存在する遺伝子が、互いに優性であるか劣性であるかは関係なく、一つが優性で他が劣性であるか、複数が優性で一つだけが劣性であるかは関係がない。
複対立遺伝の最も有名な例の一つは、人間のABO式血液型だろう。人間の血液型は血液型を決定する遺伝子座(二つ一組)に、A,B,O,の遺伝子が収まる事で決定される。
乃ち、
A-A,A-O,B-B,B-O,O-O,A-B
という、六種類の接合が有り得る。
A-A接合の遺伝子を持つ個体は、既に両方の座がAで占められていて、B,Oの遺伝子は、この座にしか納まる事が出来ない遺伝子であるから、遺伝子の中にB,O,の因子を持つ事はない。他の接合の場合も同様。
そして、この時、人間の血液型では、A=B>Oという優性劣性関係が存在する。つまり、AとBはOに比べて優性であり、AとBは対等という事。そうすると、
A型 :AA,AO
B型 :BB,BO
O型 :OO
AB型:AB
という、四つの型が出来る上がる事になる。此は、A=B>Oという優性劣勢関係だから四つになるのであり、もしも違う力関係ならもっと少ないか、或いは多くの表現型が出現することになる。
(cisAB型とかはこの場合気にしなくてよいと思うけれども、血液型ではなく別の何かしらの遺伝子座の例を考える時、人間で言うcisAB型のようなのが、存在するかもしれない。可能性としての話ですが)。
コーンスネークの場合、複対立遺伝する形質は二つ知られている。
1. 模様の遺伝子:野生型(ノーマル)、ストライプ型、モトレー型 (この時、野生型>ストライプ型=モトレー型。但し表現型は更に複雑な遺伝(多因子遺伝や、相互作用により微妙に変化し幅がある)
2. 黒色色素に関わる遺伝子:野生型(ノーマル)、アルビノ(アメラニスティック)型、ウルトラ(ウルトラハイポ)型 野生型>アルビノ=ウルトラ
後者の場合、アルビノ、ウルトラ、そして中間表現型であるウルトラメルが出るのだから不完全優性な気がしないでもないが、ノーマル型が存在するので、複対立遺伝になる。
此処では例示として、コーンの柄(ノーマル、モトレー、ストライプ)の遺伝に就いて、組み合わせと確率まで記述してみる。
模様は、科学的に検証された訳ではない(学術的な意味での科学的に)が、経験上、複対立遺伝すると思われると言われ、此の指摘は90年代前半あたりから既に為されている。
模様に関わる遺伝子座を、便宜上P=Patternとする。
P(xx)
xに代入される値により模様は決定される。
代入値は、n,m,s,の三つがある。
n=normal
m=motley
s=striped
且つ、三つの要素は、n>m=sという性質を持つとされる(可能性としてはn>m≒sとも言われる。多因子遺伝などの項目を参照。余談。パターンレス(グラニット)という模様変異もあると聞くが、遺伝の仕方や詳しい事が不明なので現時点では省略する)
組み合わせは、一つの遺伝子に於ける事なので、
P(ns)=P(sn),P(ms)=P(sm),P(nm)=P(mn)
である。nが優位であるので、nを前に書くことに統一する。msでは、mを先に書く事とする。又、以下/は基本的に省略。
上記から考えると、組み合わせは、
P(nn),P(nm),P(ns),P(ss),P(mm),P(ms),
の六通りになる。
模様の表現は、n>m=sであるので、
P(nn)=normal
P(nm)=normal
P(ns)=normal
P(ss)=striped
P(mm)=motley
P(ms)=motley*striped
(記述の順番は整理する時は、n>m>sとする)
最後のP(ms)=motley*stripedは、モトレーの模様が乱れたものや、ストライプの模様が崩れた感じのものの場合、遺伝子がこうなっていると推察されるにすぎない。通常、モトレーストライプの表現型は、ややモトレーに傾きやすいようである。P(sm)とP(ms)は同一のものと考える。
ただ、明かに変じゃないか、というのが出たなら、そこから検証していなかければならないかもしれないけれども………所謂ワイドストライプは、P(ms)の結果偶発的に生ずるものではないかと思われるが、未解明。
なんにしろ、此処では、
P(nn)=normal
P(nm)=normal
P(ns)=normal
と取り扱う。P(ms)=motley*striped は中間的になるのでしょうがどんなものかよく分からないので記述しているだけのもの。
P(nn),P(nm),P(ns),P(ss),P(mm),P(ms)の組み合わせは、6*6=36
このうち重複しているものは、六つから二つを選ぶ(=重複を許さない)組み合わせ分だから、
6!/{2!(6-2)!}=(6*5*4*3*2*1)/2*1*4*3*2*1=3*5=15
よって、考えるべき通りは、36-15=21
書き出すと、
P(nn)*P(nn)、P(nn)*P(nm)、P(nn)*P(ns)、P(nn)*P(ss)、P(nn)*P(mm)、
P(nn)*P(ms)、P(ss)*P(ss)、P(ss)*P(nm)、P(ss)*P(ns)、P(ss)*P(mm)
P(ss)*P(ms)、P(mm)*P(nm)、P(mm)*P(ns)、P(mm)*P(mm)、P(mm)*P(ms)
P(nm)*P(nm)、P(nm)*P(ns)、P(nm)*P(ms)、P(ns)*P(ns)、P(ns)*P(ms)
P(ms)*P(ms)
以上21通り
以下の記述例:
P(ao)*P(ao)
=P(aa),P(ao),P(oa),P(oo)
=P(aa)25%,P(ao)50%,P(oo)25%
組み合わせが分かり易い様に途中迄はナンバーを振ってみる。
P(n1n2)*P(n3n4)
=P(n1n3),P(n1n4),P(n2n3),P(n2n4)
=P(nn)100%,
=normal100%
P(n1n2)*P(n3m1)
=P(n1n3),P(n1m1),P(n2n3),P(n2m1)
=P(nn)50%,P(nm)50%,
=normal100%
P(n1n2)*P(n3s1)
=P(n1n3),P(n1s1),P(n2n3),P(n2s1)
=P(nn)50%,P(ns)50%,
=normal100%
P(n1n2)*P(s1s2)
=P(n1s1),P(n1s2),P(n2s1),P(n2s2)
=P(ns)100%,
=normal100%
P(n1n2)*P(m1m2)
=P(n1m1),P(n1m2),P(n2m1),P(n2m2)
=P(nm)100%,
=normal100%
P(n1n2)*P(m1s1)
=P(n1m1),P(n1s1),P(n2m1),P(n2s1)
=P(nm)50%,P(ns)50%,
=normal100%
P(s1s2)*P(s3s4)
=P(s1s3),P(s1s4),P(s2s3),P(s2s4)
=P(ss)100%,
=striped100%
P(s1s2)*P(n1m1)
=P(s1n1),P(s1m1),P(s2n1),P(s2m1)
=P(ns)50%,P(sm)50%,
=normal50%,motley*striped50%
P(s1s2)*P(n1s3)
=P(s1n1),P(s1s3),P(s2n1),P(s2s3)
=P(ns)50%,P(ss)50%,
=normal50%,striped50%
P(s1s2)*P(m1m2)
=P(s1m1),P(s1m2),P(s2m1),P(s2m2)
=P(sm)100%,
=motley*striped100%
P(s1s2)*P(m1s3)
=P(s1m1),P(s1s3),P(s2m1),P(s2s3)
=P(sm)50%,P(ss)50%,
=motley*striped50%,striped50%,
P(m1m2)*P(n1m3)
=P(m1n1),P(m1m3),P(m2n1),P(m2m3)
=P(nm)50%,P(mm)50%,
=normal50%,motley50%
P(m1m2)*P(n1s1)
=P(m1n1),P(m1s1),P(m2n1),P(m2s1)
=P(nm)50%,P(ms)50%,
=normal50%,motley*striped50%
P(m1m2)*P(m3m4)
=P(m1m3),P(m1m4),P(m2m3),P(m2m3)
=P(mm)100%,
=motley100%
P(m1m2)*P(m3s1)
=P(m1m3),P(m1s1),P(m2m3),P(m2s1)
=P(mm)50%,P(ms)50%,
=motley50%,motley*striped50%
P(n1m1)*P(n2m2)
=P(n1n2),P(n1m2),P(m1n2),P(m1m2)
=P(nn)25%,P(mm)25%,P(nm)50%,
=normal75%,motley25%
P(n1m1)*P(n2s1)
=P(n1n2),P(n1s1),P(m1n2),P(m1s1)
=P(nn)25%,P(ns)25%,P(nm)25%,P(ms)25%,
=normal75%,motley*striped25%
P(n1m1)*P(m2s1)
=P(n1m2),P(n1s1),P(m1m2),P(m1s1)=
P(nm)25%,P(ns)25%,P(mm)25%,P(ms)25%,
=normal50%,motley*striped25%,motley25%
P(n1s1)*P(n2s2)
=P(n1n2),P(n1s2),P(s1n2),P(s1s2)
=P(nn)25%,P(ns)50%,P(ss)25%,
=normal75%,striped25%
P(n1s1)*P(m1s2)
=P(n1m1),P(n1s2),P(s1m1),P(s1s2)
=P(nm)25%,P(ns)25%,P(sm)25%,P(ss)25%,
=normal50%,motley*striped25%,striped25%
P(m1s1)*P(m2s2)
=P(m1m2),P(m1s2),P(s1m2),P(s1s2)
=P(mm)25%,P(sm)50%,P(ss)25%,
=motley25%,motley*striped50%,striped25%
共優勢遺伝/co-dominant inheritance
共優勢とは、一つの座に於ける二つの遺伝子―――遺伝子Aと、遺伝子Bの関係を意味する言葉である。野生型のNormal遺伝子が優性遺伝子で、Amelanistic遺伝子が劣性遺伝子と呼ばれるように、二つの遺伝子間での力関係を指し示す。
共に支配する状態。つまり、遺伝子Aと遺伝子Bがヘテロ接合となっているときに、表現型が両者の中間となっている状態を、共優勢遺伝形質と呼称する。
劣性である遺伝子がなく、一種類の遺伝子しか存在しない座に於いては、その座の遺伝子は優性遺伝子であると呼ばれたりしないように、共優勢という言葉も二つ以上の遺伝子の関係を意味する言葉である。一つの遺伝子をして、共優勢遺伝子であると言った場合、それは、Normal遺伝子に対して共優勢の遺伝子である、という意味合いになるだろう。
優性遺伝子にあっては、ヘテロ接合であるか、ホモ接合であるかは、表現型に関与しない。
例えば、Normal遺伝子を持つ個体と、Amelanistic因子をヘテロ接合で持つ個体の場合、Amelanistic遺伝子よりも、Normal遺伝子が優性(Dominace)である為、外見上はNormal個体になる。そして、Normal遺伝子がヘテロである場合と、ホモである場合は、機能的に同じであり表現型も同一になる。
こうした遺伝子は、ホモ接合ではその遺伝子本来の機能を十全に発揮する為、ヘテロ接合の個体とはまったく異なった表現型を示す。正確には、ホモ状態での表現型が、この遺伝子が本来持っている機能を映し出したもので、ヘテロ接合時の表現型(共優勢遺伝表現型)は、そのハーフ表現形質とでも呼ぶべきものになる(同じ座に対立して存在するのがノーマル遺伝子である場合で、もしも同じ座に対立するノーマルとは異なる遺伝子があるならば、また異なったことになるだろうが………)。
一般にすぐ思い至るのは、ボールパイソンのパステル形質だが、一般的にパステルと呼ばれるのが共優勢遺伝形質で、スーパーパステルと呼ばれるのがホモ表現である。Amelanistic遺伝子の場合、ホモがAmelanistic形質で、ヘテロはヘテロAmelanisticになることを考えると、パステルという遺伝子があり、そのホモ接合がパステルで、そのヘテロ接合がハーフパステルであるという考え方も出来る。が、既にパステルとスーパーパステルという呼び方は定着しているので、今後も共優勢遺伝形質がベースであり、その上にスーパー○○がある、という風な解釈になるのだろう。どちらにしろ呼び方が変わるだけで、本質的には何も変わらないのだが(つまり、基準点を何処に置くかという相対的な違いでしかなく、どちらも等しく正しいということ)。
ex)
パステル形質は、Pastel=P、Normal=Nという遺伝子の接合である、P-Nという存在。此に対し、P-Pという接合の時、パステルはよりその形質を強く出し、スーパーパステルと呼ばれる表現型になる。
・パステル×パステル
P1N1×P2N2=P1P2(25%),P1N2(25%),N1P2(25%),N1N2(25%) が、それぞれ25%ずつ。つまり、スーパーパステルが25%、パステルが50%、ノーマルが25%で産まれる訳です。
ナミヘビ全体で見ると、幾つかそうした遺伝子は存在しているようではあるけれども、コーンに於いては、まだ確認されていないと思われる。
優性遺伝/dominant inheritance
優性という表現は、現在では顕性遺伝とすべきという動きもある。
それはさておき、ハープタイルホビーの分野で注目される遺伝子変異は、野生型に対し、劣性であるものが多かった。近年、共優勢であるものも見つかっているが、野生型に対して、優性であるものは殆ど知られていなかった(カラーミューテーションに限っているからだろうが)。
が、ボールパイソンでは優性遺伝形質が見つかっている。スパイダーと呼ばれるこの変異は、野生型に対して優性で遺伝する。ところで、此の場合の優性遺伝というのは、どういう風に遺伝するのか。
端的に言ってしまえば、スパイダー遺伝子を此迄の野生型遺伝子の位置におき、野生型遺伝子をアルビノ遺伝子などの位置に持ってくればよい。
スパイダー(ホモ)×ノーマルの子供は、言うなれば"ヘテロ・ノーマル"というものになる。何故なら、その表現型はすべてスパイダーであるからだ。そして、ホモのスパイダーと、ヘテロのスパイダーは、外見からは見分けが出来ない(何かしら他の要因も混ざれば見分けられるかもしれないが)。
此らのヘテロ・ノーマルのスパイダー同士を交配させると、ホモのスパイダーが1/4、ヘテロノーマルのスパイダーが1/2、ノーマルが1/4の確率で出てくる事になるだろう。
ところで、スパイダーはヘテロとホモで値段が違うのか、と聞いてみたところ、あまり値段差はついていないらしい。スパイダーの値段は、どちらかというとスパイダー模様の綺麗さに関わっていて、綺麗なスパイダーは高く、ちょっと微妙なスパイダーは安いそうだ。ちょっと考えると、ホモの方がお買い得な気がするのだが、たぶん、ホモの方が綺麗に出るだろうから、綺麗な個体ほどホモの可能性が高いと皆思っているのかもしれない。
そういえば、スパイダー以外にも優性遺伝する形質ってありましたね。
でも、ナミヘビでは知られていないような気がするのに、何故此処でこんな話を書くのか?と思われた方もいるかもしれませんが、ナミヘビでもいるかもしれないと思っているのです。それは、まぁ、まだ検証中なのですが………まぁ、ほら、アレのストライプがそうじゃないかな、と?(ピンと来る日とは此処で来ましたか?)
その他の遺伝
細胞質遺伝/cytoplasmic inheritance
細胞質遺伝とは、核に存在する遺伝子ではなく、細胞質に存在する遺伝因子によって起こる遺伝のこと。例えば、ミトコンドリアDNAが細胞質遺伝。
通常、遺伝の多くは、細胞核に存在する染色体に依存している。此に対し、ミトコンドリアや葉緑体など、細胞小器官に由来する遺伝があり、此らを総称して細胞質遺伝と言う。細胞小器官は細胞核とは別に、独自に遺伝構造を持ち、細胞核とは異なった遺伝方式で遺伝する。
故に、其の形質遺伝はメンデルの法則に従わない。有名な話であるが、ミトコンドリアのDNAは、動物に於いては母親から(100%ではないが、極めて高いほぼ100%と言ってよい確率で)受け継がれる。葉緑体も同じ。
果たして、この遺伝方式が関与するハープタイル・ホビーのミュータントが存在するかは分からない。
伴性遺伝/sex-linked inheritance
伴性遺伝とは、常染色体ではない染色体、乃ち、性染色体に依存する遺伝のことを言う。
性染色体自体が、或る意味伴性遺伝で、人間の場合は、Y染色体上にある遺伝子により雄個体になる。ヘビの場合は、W染色体を持つ方が雌になる。
此により、例えば、W染色体上にしか存在しない変異は、雌個体にしか遺伝しない。また、Z染色体上の変異の遺伝の場合、Z染色体を二つ持つ雄の方が雌よりも発現確率は高い。また、性染色体は通常、同一の染色体を持つ個体(ヒトならば雌、ヘビならば雄)の場合、片方の染色体が不活性化するものなので、遺伝因子を持っていても必ずしも発現しているとは限らないということを考慮に入れなくてはならない。
例としては、人間の色盲がそうであるらしい。ただ、色盲と言っても色々あり、日本人の場合、かなり高い割合で色盲を持っている。日常生活に支障が出ない場合は本人も気付いていない場合もあるという。遺伝の本には書いてなかったが、おそらく、劣性で伴性遺伝する色盲というのは全色盲のことと思われる。
余談が過ぎたが、人間の色盲はX染色体上の異常であり、かつ劣性遺伝子である。
つまり、正常な性染色体Xに対し、色盲を呈する遺伝子を持つ性染色体をX'とする。劣性に遺伝するので、もしもX染色体を二つもつ場合は、X'X'という組み合わせの時のみ色盲になり、XX'という組み合わせでは、色盲に成らない。
男性の場合、X染色体を一つしか持たない為、X'YかXYの組み合わせしかない。
乃ち、母親が色盲であるX'X'と、色盲でない父親XYの間に産まれる子供は、女の子の場合はX'Xという組み合わせであり、色盲にはならない(ヘテロであると言える)。此に対し、男の子の場合は、母親から受け継がれる性染色体は必ずX'であり、父親からはY染色体しか受け継げないので(Y染色体を受け継がなかったら、そもそも男の子にならないのだから)、必ず色盲になる。
他にも、血友病であるとかもそうであるし、人間以外の例を挙げるならばキイロショウジョウバエの白眼が伴性遺伝なのだとか。此の手の遺伝は結構あり得る気がするが、現時点で、爬虫類の突然変異の表現型で、伴性遺伝するもの、というのを聞いた事はない。
致死遺伝子/lethal gene
遺伝子には、発生から誕生、成長、成熟に到るまでの長い期間、如何に体内の特定の細胞や組織、器官が働くべきかが記されているという。但し、例えばどの細胞がどういう器官になるのか、決めている決定的な要素は分かっていないのだとか。
斯うした肉体の形成、変化を促す部分が突然変異を起こし、その作用が異常になった時、場合によってはその個体を直接的に死に至らしめる場合がある。
そのような変異を持った遺伝子は、致死遺伝子と呼称されるが、
・配偶子致死遺伝子 /gametocidal gene
・半致死遺伝子/semilethal gene
・優性致死遺伝子/dominant lethal gene
・劣性致死遺伝子/recessive lethal gene
といった幾つかの種類がある。
致死作用が発生初期に影響するものであれば発生することがなかったり、死産となるが、すべてがそうである訳ではない(例えば、肺の形成に関わる遺伝子が変異して成長に肺が形成されなくても、卵の中などで肺を介さない呼吸が行われている間は、これは致死にならない。孵化の段階ですべて死亡する)。こうした遺伝子により起こる病理的な症状を致死遺伝子症/lethal geneと言う。
そうした遺伝子を持った時点で発生自体が起こらない為、通常、我々の目に触れる事があるのは、劣性致死になる。
元々発見された経緯は、ハツカネズミの黄色の毛色のもの(通常体色:黒色に対して優性)に於いて、通常体色との雑種第一世代の分離比が、黄色:黒色で2:1になったことから、何故か、という議論になったからだとか。法則に従うならば、3:1の比率でなくてはならない。
此は結局、ハツカネズミの黄色の毛色の因子が、ホモ接合の時、致死となり、体内で死亡している為で、黄色に産まれてきているのはすべて黄色の毛の遺伝子のヘテロ結合の個体だった、という結論を見た。
此の様に、致死遺伝子の多くは優性の形質を支配する遺伝子で、ヘテロの状態で何かしらの突然変異表現型を示し、致死作用は劣性として作用する。
マウスやショウジョウバエなどで、このような遺伝子が数多く知られているが、蛇ではあまり聞かない。ボールパイソンのパイボールドという変異が、どうあっても頭部だけは白化しないのは、逆説的に言えば、頭部が白化する場合は致死になっているのでは、という推察も立つが、実際のところは不明。コーンスネークを始めとしてナミヘビで、此の様な遺伝の仕方をする変異は耳にしないが、やたら死籠もりや未発生が多い変異があったら、それは此の手の遺伝子を抱えているのかもしれない。
同義遺伝子/multiple gene
生物の、とある表現型を表す遺伝子が、複数の異なる遺伝子座に存在する場合、それらの遺伝子は同義であると言う。表面に目に見えるケースでは、それらの遺伝子がそれぞれ、その座に於いて対立遺伝子を持っていなくてはならない。此らは植物の葉の形などを決定する遺伝子に多いのだとか。
例えば、とある植物で、葉っぱが丸い形をしているものと、細長い形をしているものがあるとする。それぞれを独立に交配し、常に、丸い葉っぱのものは丸い葉っぱのもの、細長い葉っぱのものは細長い葉っぱになるようにしておく。
そして、此の二つの雑種一代目をとると、その総てが丸い葉っぱのものとなった。更に、その雑種一代目同士を交配した時、丸いものと、細長いものが、15:1に分離した。
このケースでは、葉っぱを丸くする遺伝子が、二つの異なる遺伝子座にある優性遺伝子AとBであった場合の例であり、メンデルの法則で理解することが出来る。
乃ち、遺伝子座Aに於いてAとaという遺伝子があり、遺伝子座Bに於いてはBとbという遺伝子があった。そして、AとBは異なる遺伝子座にあるが、どちらも、「葉っぱを丸くする」という意味を持つ優性の遺伝子であった。その場合、二つの遺伝子座での結合が、共に、aaとbbである場合にのみ、葉っぱは丸くならず、細長い劣性形質が現れることになる。
この場合、葉っぱが細長くなるのは、二重劣性遺伝表現型であると考える事が出来るので、その出現額率は1/16ということになる。
もしも、同義遺伝子の数が二つではなく三つ、四つとなっていき、また遺伝子の持つ性質が複数あればより複雑化することになる。
抑制遺伝子/inhibitor gene
抑制遺伝子とは、とある遺伝子の作用を抑える働きを持つ遺伝子のこと。通常、此が表面上に出てくる問題となるのは、劣性遺伝子に対して抑制機能を持つ遺伝子が存在する場合である。
グッピーの遺伝で知られる遺伝で、グッピーの尾ひれを伸ばす(燕の尾のようであるということで、スワローテイルからとって、スワロー遺伝子などと呼ばれる)
遺伝子に対して、尾が伸びるのを抑制する機能を持つ遺伝子A(antiということで便宜上名付けた)が存在する。
表現形遺伝子Sに対し、抑制する機能を持つ遺伝子Aが存在する場合、遺伝はどうなるのか。
上記例のスワローでは、表現型遺伝子Sは、ヘテロ接合であっても表現型になりうるが、同時に抑制遺伝子Aもヘテロ接合で抑制機能を発揮する。また、抑制遺伝子は一つあるだけで表現型に対し優性になっている(つまり、Sがホモ接合であっても、抑制遺伝子Aがヘテロ接合として一つあるだけで抑制機能は発揮されノーマルになる)。
抑制機能は必ずしも上記例のように作用するとは限らないと思われるので、例えば抑制遺伝子がホモ接合の時のみ機能を発揮する場合や、ホモ接合に対し抑制遺伝子がヘテロ接合であれば表現型が抑制されずに表面に出てくるケースなど、様々なパターンが存在すると思われる。それぞれにおいて発現する確率などが異なってくると言える。
此処では、先述したスワロー遺伝子を例にとってパンネットのスクエアを用いて説明する。
スワロー遺伝子と抑制遺伝子をそれぞれヘテロ接合の状態で持っている個体(外見上ノーマルになる)同士の交配を考えるとすると、遺伝子はS(n.s)A(n,a)であり、
Swallow (n=normal, s=Swallow) Anti_Swallow (n=normal, a=Anti_Swallow) |
S(n)A(n) 1/4 | S(n)A(a) 1/4 | S(s)A(n) 1/4 | S(s)A(a) 1/4 |
| S(n)A(n) 1/4 | S(nn)A(nn) 1/16 | S(nn)A(na) 1/16 | S(ns)A(nn) 1/16 | S(ns)A(na) 1/16 |
| S(n)A(a) 1/4 | S(nn)A(na) 1/16 | S(nn)A(aa) 1/16 | S(ns)A(na) 1/16 | S(ns)A(aa) 1/16 |
| S(s)A(n) 1/4 | S(ns)A(nn) 1/16 | S(ns)A(na) 1/16 | S(ss)A(nn) 1/16 | S(ss)A(na) 1/16 |
| S(s)A(a) 1/4 | S(ns)A(na) 1/16 | S(ns)A(aa) 1/16 | S(ss)A(na) 1/16 | S(ss)A(aa) 1/16 |
よって、スワローの出現確率は3/16になる。
グッピーに於けるスワローの出現はヘテロでも表現型になるため此の様な確率になっているが、大抵ヘビの遺伝変異は劣性遺伝であることを考えると、ヘビでもし此の様な遺伝方式による遺伝があるならば、出現確率は1/16になるものとなるのではないかと思われる。
或る意味、この抑制遺伝子による遺伝は、同義遺伝子による遺伝のバリエーションの一つと考える事が出来るだろうか。あちらが出現する為に複数座に於いて同じ効果を持つ遺伝子がホモ接合で揃わなければならないのに対し、此方は一方の座でホモ接合であり、もう一方の座でノーマル遺伝子によるホモ接合でなくてはならないのであり、よって出現確率は一見すれば同等になる。
但し、出てきた表現型を系統として固定していく場合、同義遺伝子であれば表現型同士を交配していけば、その子はすべてその表現型になるのに対し、抑制遺伝子による表現型は、遺伝子が劣性表現型でない場合(スワロー遺伝子のようなものである場合)、表現型同士を交配してもノーマル表現型が出てくる可能性がある。上のパンネットのスクエアで濃いピンクで囲んだ部分同士の交配の場合がそれにあたり、その場合、1/4の確率でノーマルは産まれるわけだが、此処で重要なのは、スワロー遺伝子のような変異は、ヘテロ接合でもホモ接合でも同じ表現になるため、外見から区別することはできないということである。よって、交配してノーマル表現が出るかどうかを見ないことには、ヘテロ接合なのかホモ接合なのかを見極める事は出来ない。まぁ、さほど気にする事もないかもしれないが…………
相加遺伝子/additive gene
(必要ないから書かなくて良いんではないだろーか、みたいな)
多因子遺伝/multifactorial genetic
(難しいし、別に書かなくても良いんではないだろーか)